Journal of the American Optometric Association, Why do virtually all computer operators suffer to some degree from headaches, asthenopia, fatigue, various types of head-neck-shoulder distress, as well as hand-wrist discomfort? Simply because most existing computer hardware designs have not adequately taken into account the way human beings would be functionally most comfortable in performing ocular and orthopedic near-point tasks. Rather, the individual computer hardware components developed first and have been juxtaposed in the most convenient manner, and we the users are in effect, the "squares pegs" who have had to accommodate ourself to fit the "round holes." The internal adjustment options and the external add-ons to the existing systems, (i.e., Polaroid or mesh overlay screens, various image/background colors, differential screen resolutions, luminance/contrast controls, ergonomic chairs and work stands) all contribute incrementally to the user comfort and efficiency.1,2 We are seriously disadvantaged however, because we are starting from the circumstance of poorly designed and integrated computer components. To create a more ideal working environment, a new schema has been postulated which integrates the ocular neurology and orthopedic biomechanics. Ocular neurology This would appear to be a violation of Listing's Law, which describes monocular movements only about X and Y axes, and which makes no provision for rotational movement about the Z axes. Allen and Carter so concluded this, and suggested that excyclotorsion be included as part of the near reflex.6 These cyclofusional movements have been classified as psycho-optical responses which are first initiated by attention to the target appearance, and then responded to by reflex. To the extent that there is less excyclotorsion, there will be less cyclofusional movement of the eye to restore and maintain the visual "vertical" in the interest of sustaining single binocular vision, and the by-product of this would be less expenditure of energy (and presumably less fatigue) (Table 1).
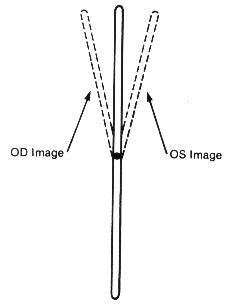
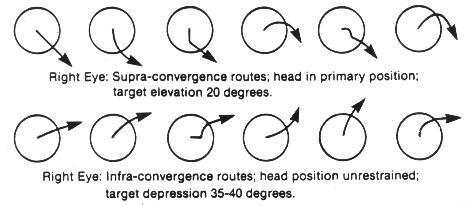
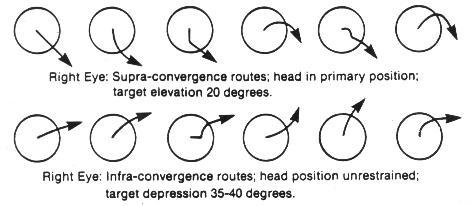
Experimentally it has been demonstrated that relatively small target variations from parallelism of 5-8% will create the subjective experience of blurring, disruption, and diplopia.7 It can often be demonstrated monocularly that there is an inward rotation of the upper end of a vertical target line as the gaze is shifted from a depressed position, through the primary and into an elevated position. In this circumstance there would be an excyclorotated retinal image, and sequentially a compensatory extraocular muscle excyclotorsion (Figure 1). This effect can also be noted monocularly with a tipping backward of the upper end of a vertical target line (Figure 2). Similarly, studies of the incomitance of heterophorias relating to elevation-of-gaze and depression-of-gaze have indicated that the predominant phoric variation is that of a "V" pattern (in which there is more exophoria associated with elevation-of-gaze, and less exophoria associated with depression-of-gaze). This effect was more pronounced with near-point fixation of 33cm, than at a 3m testing distance. The "V" pattern incidence and extent increased with larger gaze elevation/depression angles (from 20-40o).8
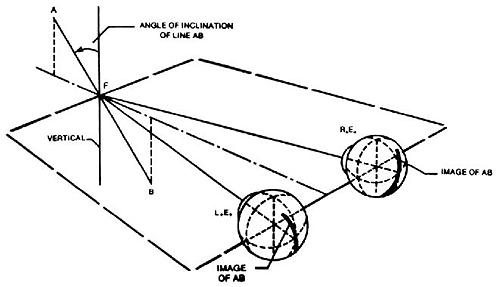
Over a long time-span, evolutionary changes occurred permitting human forebears, out of necessity, to view distant objects clearly and with some sense of depth. Duke-Elder comments that "biologically the eyes were adapted for relatively simple purposes - to look for enemies and for food."9 Large overlapping visual areas producing binocularity became prerequisite to predatory skills.10 To sustain the advantages of single binocular vision, cyclofusional ocular movements became a necessity " . . . in order that vertical objects such as walls, trees and so forth may maintain their orientation as one raises or lowers the eyes to look at various parts. . . ."7 If a valid measure of our human uniqueness is that of being tool-makers and tool-users, then it logically follows that we would have to be looking clearly at the end of our forelimb appendages to accomplish the task at hand (figuratively and literally). The side-benefit of the binocularity would have been a greater stereopsis sense at near-point because of the significant lateral displacement of the eyes in the skull (4-7cm), and the increased amount of convergence being used. While on all fours in an early quadripedal age, the linear distance from the plane of our eyes to the ground between our unflexed forelimbs was only incrementally greater than a normal reading position distance for modern man. Whether we were still herbivores breaking apart nuts, or had become omnivores and were now also picking apart the carcass of a kill, these tasks would have been easier looking downward. Once the tool manufacturing was completed, utilization still required point-of-use visual clarity (the keyboard), and point-of-resultant-tool-usage clarity (the screen), and while prior experience might partially and temporarily supplant the visual-touch mechanism, memory alone is unequal to ongoing, efficient task performance. We simply have to be looking at the things we do. In our current bipedal mode, it is obvious that we would expend less energy in tool manipulation if our forelimbs were in that position that least conflicts with gravity (a rather practical method of conserving physical strength by not bearing the weight of our forelimbs in an unsupported and elevated attitude). The result is a happy synchronicity of the most comfortable tool-using posture with depression-of-gaze.
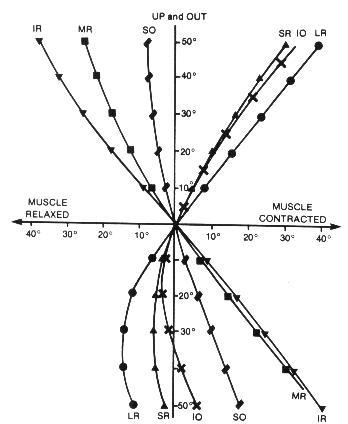
Is the proposed Near Quintad Function neurological, or is it simply that during convergence, extraocular muscle synergism provides greater action by the combination of those muscles resulting in a depressed position of gaze and consequently less excyclotorsion? It has been established that the closeness of the insertion of the extraocular muscle to the limbus, and the greatness of its muscle mass will determine its relative activity during contraction. The medial rectus ranks first in both categories, and the inferior rectus has the next closest insertion to the limbus, and the third widest tendon diameter (insertion spiral of Tilleau).11 Further from Duke-Elder, "Thus convergence, so constantly required for near work, with the exception of its presence to slight degrees in the higher apes, is a new function unique to man and is frequently one of the first to break down in the conditions of strain."9 Many researchers have studied and detailed differing conclusions regarding binocular extraocular muscle function, but the consensus is that: Any position of gaze is the sum total of cooperation of all the extraocular muscles, and that any schematic model of individual muscle action would have to be considered hypothetical. There are different sets of muscle planes and axes of rotation for each position of gaze because the insertions of, and the states of contraction of the extraocular muscles are changed for each position of gaze.12 And what is contradictory is that: "In the primary position, the axis of the horizontal muscle pair is almost precisely vertical.... In extreme adduction, however, the axis inclines about 8o... with the result that there are small components of depression and excycloduction."12 Solomons has shown the variations of extraocular muscle action for various positions of gaze and comments (with regard to primary muscle actions) that "... In the horizontal meridian, the lateral rectus abducts and the medial rectus adducts; however they both elevate the eye very slightly, with the lateral rectus being slightly more effective." However, while detailing secondary and tertiary actions of the extraocular muscles it becomes apparent that each individual muscle contributes differential contractive and torsive efforts to the resultant ocular position. With adduction, only the inferior rectus and superior oblique exert depressive effort. With depression, there is now synergistic effort by four muscles to effect/sustain this position: medial rectus, inferior rectus, superior oblique, lateral rectus. With depression/adduction, there are five muscles which contribute (the superior rectus now adds to adduction). Further, according to Solomons, " ... When the eye looks 'down and in,' the inferior rectus is the strongest acting muscle, aided almost equally by the medial rectus, and to a lesser extent by the superior oblique. For movements greater than about 35o the inferior oblique also becomes involved"13 (Figure 3). There have been animal studies in which the extraocular muscles in monkeys have been severed and the insertions transposed (medial rectus/lateral rectus), and where the superior oblique muscle insertion has been substituted for a rectus muscle insertion. Postsurgically within days, the researchers found the ocular motility of the eye to have returned to its preoperative condition.14 This suggests other nerve pathways of stimulation-response which influence ocular movements. Despite these observational differences, what is implied is that accommodation/convergence/miosis are in some manner linked with depression-of-gaze and excyclotorsion. According to Birnbaum, "Although efficient near-point function requires precise matching between accommodation and convergence, a tendency for convergence to localize closer than accommodation is intrinsic to the near-point demands imposed by our culture. The high level of psychological stress pervasive in our society is an additional factor generating sympathetic activation and overconvergence.15 The convergence needed to perform near-work tasks at a 40cm distance, with an assumed average interpupillary distance of 63mm, results in angular deviation from parallelism of 12o. Shortening the working distance to 20cm doubles the angular deviation to 24o. Typically though, accommodation does lag behind convergence resulting in an average of 4.2 diopters of accommodation for each prism diopter of convergence (as opposed to an ideal geometric match of 6 diopters: 1 prism diopter). What then is the mechanism to ensure near-point target clarity? With ocular synkinesis (The Near Triad), although there is intermediation between the component functions, it is the disparate images that initiate vergences first, which then affects the state of accommodation (in seeking a clear image at the new "verged" position), which then causes pupillary miosis (resulting in an increased depth of focus), which then lessens some of the accommodative effort required for a clear target image. Further, "The pupil is known to constrict with sleepiness, boredom, or fatigue and, if constricted, would increase the depth of focus, reducing the requirement for accommodation."16 Therefore, to whatever near-point the eyes are accommodated for, we are overconverged upon a yet closer near-point, and there would be increased excylotorsion.
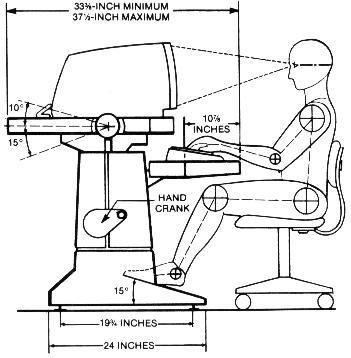
Borish has stated: " . . . The eye is seldom turned 10-12o, as head movements complete the needed angular turn for most fixations. In theory the movement possible is only 1/8 or 1/9 of the total."17 Von Noorden supports this view that measurements of eye positions exceeding 15o from the primary have little significance in our functional visual lives, and cites other research that horizontal ocular excursions were minimal (6-8o) for most visual functions. Generally, ocular movements preceded head movements, and the head movements were greater with increased difficulty of the near-work task.5 The simplest way to demonstrate the depression-of-gaze effect is to conduct a push-up-convergence test by having the subject in a standing or seated position with a primary head position, and then introducing a small, finite target brought directly inward along the midline of the primary horizontal plane of the two eyes. The subject's eyes will at some point during converging, be in a depressed gaze position, and will achieve this position by various routes. Each eye may also travel a different path while converging. If this finite target is introduced from above the primary horizontal, what is observed is a converging-inward, sometimes upward-arcuate-downward ocular movement, which always ends in a lowered ocular position. When the finite target is introduced from below the primary horizontal (and permitting the subject to drop the head forward and downward into whatever position is most natural), what is observed is principally a converging movement with a slight upward component that can occur at the beginning, midpoint, or toward the end of the sequence (Figure 4). This author has observed and detailed push-up-convergence responses of 100 individuals (all ages, both sexes, all refractive errors) for three positions. The finite target was moved inward along the midline from 60cm to 20cm. In the primary and supra positions, the head position was restrained in an upright posture because otherwise, the subject would have tended to elevate the chin and tip the head backward slightly (capitas extension) as the target approached, and this would have produced a false appearance of convergence without a depression-of-gaze component. Capitas extension is noted in pre-presbyopes and is exaggerated in over 40 operators who gradually need supplemental presbyopic corrections to see clearly the screen/keyboard/copy-viewing area, and because supraduction ability is also lessening. If screens were placed at least 35-40o below the primary horizontal plane, and angled backward 45o to the vertical, then the wearing of conventional multifocal spectacle corrections would eliminate the need for this orthopedic imposture.
In virtually all the primary-convergence observations, a depression-of-gaze component was noted. With infra-testing, the head position was left free so that the target-viewing was the combination of both head flexion and ocular depression-of-gaze. What this illustrates is that, either through volition or involition (or both), the eyes will migrate to a depressed position-of-gaze during the adductive process. Further, virtually every subject notices less subjective comfort in trying to converge upon an incoming target from a straight ahead or elevated position. The greatest subjective degree of comfort occurs when convergence and depression-of-gaze are combined. That supraduction function, diminishing from an average capability of 40o (patients age 5-14) to 16o (patients age 85-94),18 supports the depression-of-gaze/maximum comfort thesis. Occasionally with conventional computer usage, a capitas-flexion position is noted which creates a false elevation-of-gaze. It is probable that this operator has opted to bear with whatever distress the increased supraduction and excyclotorsion will produce, and to avoid capitas-extension postural difficulties (Figure 5). IBM Human Factors Center states that "a comfortable line of sight is 10o or more below the horizontal." (My observation is that this is only partially correct.) "Hence, the display should be located so that the normal line of vision falls in the upper half of the display."19 (My observation is that this is stressful to the user.) The National Research Council cites "a preferred downward slope of the line of sight from 15-30o."16 (My observation is that this is still inadequate.) This compromise of good ocular function was probably accepted as means of minimizing reflected glare that would have accompanied the sufficient lowering and angling backward of the screen. With existing computer usage, we are utilizing relatively large saccades (30-45o) to continually reposition the screen/keyboard/copy-viewing-area targets onto the fovea. With large angular-difference refixations, transient dysmetria (under-shooting the subsequent fixation point) is not uncommon.18
There is unavoidable perceptual disorientation of visual space created by torsional imbalances when the eyes are in a tertiary position.10 This condition is persistive, because astigmias corrected with a spectacle Rx or hydrophilic contact lenses are based upon examination techniques derived while the head and eyes are in a primary position. Rarely do we encounter astigmias which are totally corneal and with no residual component, so that the refractive error is fully corrected with a rigid contact lens prosthetic (in this instance, torsional imbalances would not be exacerbated by working in a skewed position). Therefore copy-viewing areas placed laterally and inferiorly to typewriters and computers are poor. Ideally the copy-viewing area should be along the visual mid-line and at least 35-40o below the primary horizontal plane (Figure 6). Although field-of-fixation (an area within which fixation can be accomplished by moving only the eye and not the head) is clinically demonstrated to be relatively large (40-50o), it is less binocularly than monocularly, and its amplitude diminishes with age. The presence of a stiff neck reveals the subjective difficulty in executing ocular, sans head movements.5 The pursuit system which matches eye velocity to target velocity, in viewing a 7" x 9" screen from an 18" working distance, will have to encompass an approximate angular range of 30+o vertically and 40+o horizontally. Following the advancing cursor as data is inputted, appears to be well within that speed that would detract from the comprehension of the operator. Vertical scrolling speed however, appears to exceed comprehension levels, and it is uncertain whether vertical scanning capability is being detracted from by successively appearing targets. References
Reproduced from
the November 1987 issue of the Journal of the American Optometric
Association
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||